
Measuring the yield of Cannabis sativa as a response to either automated irrigation systems or traditional hand watering methods in a controlled agriculture environment.
NOTE: If you are interested in supporting KIS Organics and seeing more of this research or want help utilizing a data-driven Blumat design in your garden, you can click here for help with a custom Blumat design.
Ben Higgins†§ and Tad Hussey†
§ - Gold Leaf Gardens, Lacey WA
† - KIS Organics, Redmond WA
Corresponding author: Ben Higgins – ben@kisorganics.com
Tad Hussey: tad@kisorganics.com
Abstract:
Background
The cultivation of Cannabis sativa is expanding all over the globe. However, the existence of differing laws and regulations have made it challenging to study the plant. Given the lack of scientific inquiry into C. sativa, cultivation facilities are likely not expressing neither the chemical nor phenotypic maxima of its genetics, thus creating a yield gap. One variable that would likely assist in closing this yield gap is irrigation. Soil moisture is known to play a critical role in shaping physiological and biological responses in both plants and the microbial communities within the soil. The present study developed two hypotheses to investigate the relationship between soil moisture content stability (a variable untested in C. sativa) and yield (aboveground biomass [AGB]). A total of 112 C. sativa plants from five distinct cultivars were grown for 73 days until harvested. The difference between the AGB of plants grown in experimental beds (n=5) which were irrigated with Blumat systems that automatically sense soil water potential were compared to control beds which were irrigated by hand (n=8) and analyzed using an analysis of variance (ANOVA). To determine the variation in soil moisture content throughout a two week period, a digital tensiometer was used to measure the water potential for all beds. These data were then analyzed using a Wilcoxon Rank-Sum test, followed by a Levene’s test for equal variance. The results show that when grown in living soil with stable water pressures, four of five cultivars tested increased their AGB by 20-29%, with only a single cultivar decreasing (-10%) in AGB. Cumulatlively, cultivars grown in living soil beds fitted with Blumat irrigation systems were significicantly larger, averaging 24.7% more AGB (x̄ = 5.08 lbs. per 8 plant trellis) than did plants grown in identical medium irrigated by hand (x̄ = 3.8 lbs. per 8 plant trellis). Additionally, living soil beds that were fitted with an automated irrigation system that relied upon water potential held a significantly more narrow water pressure range relative to beds which were irrigated by hand. Irrigation is a key variable to consider when attempting to further close the yield gap for C. sativa. To maximize the yield of numerous cultivars grown simultaneously, commercial facilities should consider utilizing automated irrigation systems that have some degree of interaction with the soil rather than relying on timers to determine when to start/stop irrigation cycles.
Introduction:
The laws governing the cultivation and consumption of Cannabis sativa are changing all over the globe. However, the rate at which various countries or other jurisdictions (e.g., states, counties, etc.) legalize its usage and purpose varies greatly. Canada, for example, has passed laws resulting in the legalization of both medicinal and recreational on a federal level. However, to the south, the United States has left this decision to the individual states, rendering C. sativa still listed as a Schedule 1 drug of the Controlled Substances Act. However, as of July 2020, 11 states have legalized both medicinal and recreational usage, whereas an additional 31 states have at least some usage (e.g., medicinal, CBD oil usage only, etc.), leaving only 8 states in the union where C. sativa is still illegal (https://disa.com/map-of-marijuana-legality-by-state). The decision to leave C. sativa federally listed as an illegal drug has vast implications not only relating to both the medicinal and recreational usage of C. sativa, but also to the cultivation and basic understanding of the plant as a whole.
In the United States, the two most common avenues used to pursue research and advance our understanding, are our governmental and academic institutions. However, because of its federal listing as a controlled substance these two pathways have remained largely closed, resulting in a vast lacuna of scientific research and inquiry. Research institutions that would typically serve as the venue for hypothesis testing in laboratory settings are unavailable to the emerging scientific community focusing on all aspects of the biology of C. sativa. Instead, most of this work originates from either home grows or commercial production facilities, however both of these options present challenges when attempting to conduct scientific research (e.g., laboratory level variable control, conflicts between data collection and standard operating procedures, etc.). As a result, the impacts that the psychoactive cannabinoid tetrahydrocannabinol (THC), along with over 200 terpenes, cannabinoids, and additional phenolic acids (Andre et al. 2016) have on the human body remain mixed, with both positive and negative results reported (Hill et al. 2012; Giacoppo et al. 2014; Volkow et al. 2014; Burstein 2015; Backer et al. 2019). In addition to the lack of research on how C. sativa and its myriad of secondary metabolites impact human physiology, very little is understood about how to cultivate and propagate this plant on a commercial scale to its phenotypic and genetic potential. This point was made clear from a meta-analysis which focused on the attempt to close the yield gap for C. sativa (see Backer et al. 2019). The authors review a host of variables that were investigated in relation to the impact of production methods on yield and cannabinoid profiles, such as the usage of microbial inoculants (Winston et al. 2014, light intensity and photoperiod (Chandra et al. 2011a, 2015), temperature (Chandra et al. 2011b), and other physiological stresses (see Lyndon et al. 1987; Marti et al. 2014). In short, the authors highlight that at present, the projected annual production of C. sativa on a commercial scale is based not on yield per square meter, but largely on facility size (Backer et al. 2019). Thus, most current cultivation methodologies are not accessing the vast genetic diversity of C. sativa, and are therefore limiting the maximum phenotypic expressions (e.g., crop yield and cannabinoid profiles) of C. sativa grown on a commercial scale. It is important to note that while robust, the recent aforementioned meta-analysis does not highlight any studies that comment on how irrigation rates and the resultantant soil moisture content may help close the yield gap - a variable known to be critical in a host of variables that range from plant health, to overall plant survival.
The water content in soils affects both the physiological state of microorganisms and plants (Walker et al. 2003; Borowik and Wyszkowska 2016). For example, respiration depends more heavily on soil moisture content than on temperature (Silva et al. 2008). Properly saturated soils sustain more functionally diverse microbial communities, thus allowing for increases in the cycling of a broader range of nutrients (Williams and Rice 2007; Borowik and Wyszkowska 2016). However, large deviations from well-moist soils can result in dramatic shifts in the ecology of soil microbiology as is observed in excessive soil moisture levels (i.e., flooding), and the associated decline in microorganism biomass (Silva et al. 2008; Unger et al. 2009). Similarly, drought can also disturb soil homeostasis, thus altering the biological community profile within the soil and its trophic structure (Kim et al. 2008). Therefore, to further attempt to close the yield gap, holding the soil moisture content stable throughout the duration of the grow cycle woud likely provide the vast microbiology associated with living soils a favourable environment.
The objective of the present study was to determine the role that a constant soil moisture content had on plant growth (i.e., aboveground biomass [AGB]) relative to typical hand-watering practices. Two hypotheses were formulated relating to the relationship between soil moisture content stability and C. sativa growth. The first hypothesis tested if C. sativa grown in living soils with a constant soil moisture content via irrigation through an automated irrigation system would produce larger plants across a range of genotypes relative to those that are grown in identical medium that were irrigated by hand. The second hypothesis was designed to test if an automated irrigation system that functioned not on timers, but rather on soil water potential pressures, would create a more stable soil environment relative to standard hand watering practices.
Methods:
Plant Growth
A total of 112 C. sativa plants spanning five phenotypes were grown indoors under controlled conditions at Gold Leaf Gardens, a commercial production facility in Lacey, WA. Throughout the entirety of the experiment, plants were grown in homogenous beds (n=13) filled with KIS Organics Biochar soil mix medium. Initially, plants were allowed to grow for 10 days in #1 containers under 1000-Watt Double-Ended HPS light fixtures with a 24h photoperiod. Throughout this vegetative stage, all plants were fed and watered using Gold Leaf Gardens’ proprietary input recipes. After their initial vegetative stage, plants were then transferred into homogenous living-soil beds (n=8 plants per bed) where they were then allowed to continue to vegetate for an additional 10 days (via 24h photoperiod), prior to inducing them to flower by adjusting the photoperiod of the flower room. Plants were then allowed to flower for an additional 60 days prior to harvesting. Throughout the entirety of the grow cycle, all plants were manually defoliated as is standard for the operating procedures of the grow facility. Importantly, all plants tested in the current study were carefully defoliated during the two days prior to harvesting as to reduce the amount of AGB that could be contributed to leaf number and size, which would insert error during the final weighing.
Experimental design
Five cultivars were randomly selected for this experiment (Birthday Cake; n=32, Pineapple Mimosa; n=16, Animal Sherbert #2; n=24, Wedding Cake; n=24, and Mimosa; n=16). All environmental parameters were held constant throughout the final 10 days of vegetative growth, and for the 7 weeks of flowering, including the same water source. The flower room was supplemented with 1200 (+/- 100) ppm of CO2 throughout the entirety of the grow. All clones were transferred into the flower room at the same time, and were randomly selected to be placed into the beds using a random number generator to control for transplant bias. A minimum of 16 plants were selected for experimentation, resulting in one full bed being irrigated using Blumats systems, and the other bed being watered by hand. It was not possible to place treatments side by side because of constraints within the routing of the Blumats systems water supply line, however cultivars were placed opposite one another across the aisle to attempt to mediate exposures to different light intensities (Fig. 1) and/or the potential for the development of microclimates within the flower room.

Fig. 1: Experimental setup of beds irrigated autonomously using Blumat Systems (left row) and identical cultivars of C. sativa plants watered using traditional hand watering methods (right row). Note that beds fitted with Blumat systems received trellis support prior to beds that were irrigated by hand.
Blumat systems were designed to utilize Blusoak driptape as the means for water delivery (Fig. 2). This system used constant positive head pressure to drive water delivery to the soil. The rate of water delivery is determined by moisture sensors placed within the soil (n=2 per 3’x4’x1’ bed). Thus, when moisture content lowers, the sensors allow water to flow through the sensor and into the driptape, irrigating the soil. Once the osmotic pressure in the soil increases past that within the sensor, the differential in water potential refills the sensor, increasing the internal pressure and thus closing the valve. All sensors were set up to maintain the soil at ~100 millibar (mbar) throughout the entirety of the grow cycle. This value was chosen as it has previously been documented to benefit soil microbial biological communities (Palmer and Hussey, unpublished data). Conversely, beds not equipped with the Blumat systems were watered ad libitum by gardeners (n=4) with no less than 10 years of cultivating C. sativa using these methods.

Fig. 2: Detail of Blumat and Blusoak driptape irrigation layout. Red tubing is the water supply line, which enters the Blumat sensors (within blue circles). The white tube that winds around the plants is the Blusoak driptape. The driptape is staked down onto the soil roughly every 4 ft.
To gauge soil moisture level maintenance relative to hand-watered methods, soil moisture content (i.e., water potential) was measured for 14 consecutive days between 11 March, 2019, and 24 March, 2019 for all experimental beds (n=13) using a digital tensiometer. The soil moisture of each bed was measured by inserting the tensiometer exactly 5” into the soil at a random location. The tensiometer was then allowed to acclimate to the soil for 30 minutes prior to measuring the moisture content in millibar (mbar).
At the end of their flowering cycle (63 days), all plants were cut down at the base of their stalk (i.e., at the intersection of the stalk and the soil). Trellises were then inverted, containing the eight plants within the gardening mesh. The trellis legs were removed, and the entire structure was then weighed using a digital pesola scale (Fig. 3). Repeated measurements of the empty trellis with gardening mesh resulted in a tare weight of 3 lbs. Therefore, 3 lbs. were subtracted from all trellises and the remaining weight was recorded. This value was therefore the summation of the aboveground biomass (AGB) of eight plant batches. Dry weights of the harvested plants were not analyzed, as these data could not be collected because of the workflow design of the processing facility.

Fig. 3: Data collection for aboveground biomass (i.e., wet weight) using a digital pesola scale (+/- 0.01 lbs). Trellis weights were tared, and subtracted from final weights.
Statistical analysis
All statistics were carried out in R 3.6.2 (R Core Team 2019). An analysis of variance (ANOVA) was used to test the difference between the two irrigation treatments. All data passed Levene’s test for equal variance prior to ANOVA analyses. A Wilcoxon Rank-Sum test was used to analyze the difference in water potential between the two treatments. Variation in soil moisture content between the two treatments were also analyzed using Levene’s test for equal variance.
Results:
C. sativa yield in relation to irrigation method
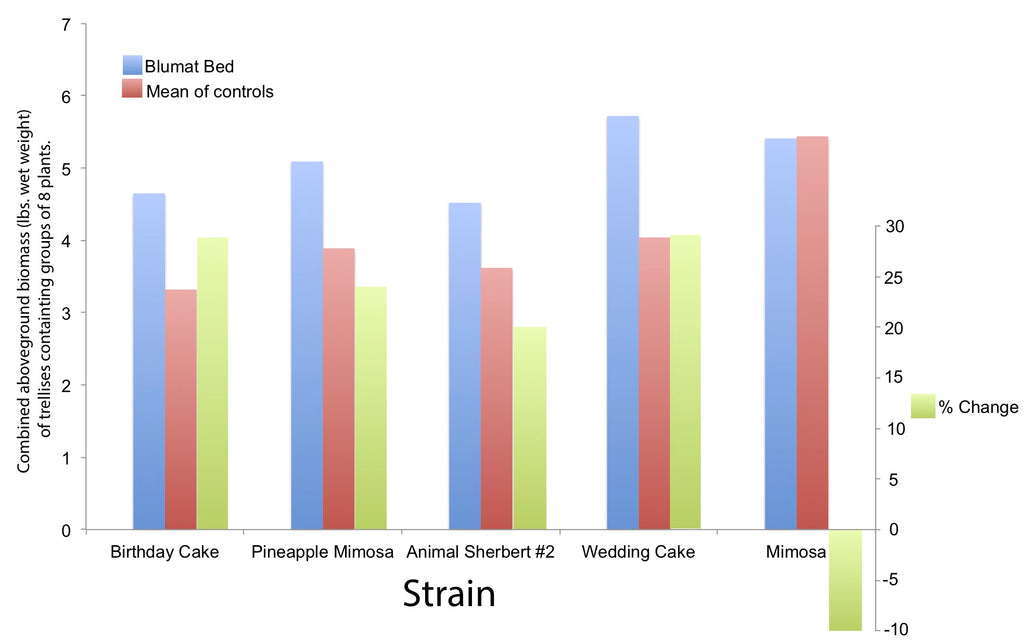
Overall, the cultivars selected for the experiment performed better when irrigated using Blumat systems, relative to those that were irrigated by hand. Objectively, plants were taller earlier, and flower mass also appeared to be larger (Fig. 4), resulting in gardeners installing trellises for plants that were irrigated with Blumats sytems a week earlier than those that were watered by hand (see Fig. 1). However, this signal was not uniform across all cultivars, as Mimosa was the only cultivar that performed better under hand water irrigation (Fig. 5). The remaining four cultivars did produce more AGB when irrigated with Blumat systems. Both Birthday Cake and Wedding Cake cultivars showed an increase of 29% in AGB at the time of harvest, whereas Pineapple Mimosa and Animal Sherbet #2 increased their yields by 24% and 20%, respectively (Fig. 6). When the treatments were combined, plants that were grown utilizing Blumat systems produced significantly more AGB than those that were hand watered (ANOVA, F10.16, d.f. = 11, P < 0.01; Fig. 7). On average, Blumat systems beds yielded 24.7% more AGB (mean = 5.08 lbs. per 8 plant trellis), relative to those that were hand watered yielded (mean = 3.8 lbs. per 8 plant trellis).

Fig. 4: Day 60 of flower of experimental trial. Note that beds containing Blumat irrigation systems are on the right, and beds irrigated by hand are on the left.

Fig. 5: Comparison of groups of eight C. sativa plants grown in homogenous living soil beds. Dark bars represent the AGB of plants grown with Blumat Systems. Additional bars indicated control bed(s). The number of controls varied between strains because of space constraints within the grow room.

Fig. 6: Total comparisons of C. sativa cultivars grown in beds fitted with Blumat Systems vs. hand-watered beds. Left y-axis corresponds to the data for blumat and mean of control beds (blue and red bars). Green bars represent the % change and correspond to the right y-axis.

Fig. 7: Mean aboveground biomass (AGB) of 8 plant trellises grown under different irrigation methods. Hand watered beds averaged 3.8 lbs. per trellis (n=8 beds), whereas beds that were irrigated with Blumat systems averaged 5.1 lbs. per trellis (n=5 beds).
Soil moisture consistency
Over the course of the two-week trials of sustained water potential measurements, the soil moisture content of living soil beds that were irrigated with Blumat systems remained much more stable than those that were hand watered (Fig. 8). Throughout the five beds that were irrigated with Blumat systems, the mean water potential was 94 mbar, with the most extreme reading at 141 mbar, and a minimum measurement of 42 mbar. For the hand watered beds, the mean water potential was 114 mbar, with a maximum value of 211 mbar, and a minimum reading of 39 mbar. Thus, hand watered beds were further from the target water potential value of 100 mbar (+14 vs. -6 for Blumat systems beds), and had both the highest, and lowest readings. Additionally, the variation in water potential within the hand watered beds were significantly higher than those beds irrigated with Blumat systems (Wilcoxon Rank-Sum test, W8655.5, P < 0.001; Fig. 9).

Fig. 8: Mean soil moisture readings of all experimental beds for 14 consecutive days. Red data series represents the beds irrigated by hand watering, while blue data series indicates beds irrigated by Blumat systems. Black dashed line at 100 mbar indicate the target soil moisture content based on soil mix structure and drainage. Error bars for each data point are +/- 1 S.D. of the mean.

Fig. 9: Variation of water potential readings across living soil beds irrigated by two different methods. Over the same period of time, beds irrigated with Blumat systems had significantly less variation in soil moisture content.
Discussion
To our knowledge, the present study provides the first description of C. sativa in relation to irrigation methods and soil moisture content while grown in an indoor production facility. The results of this study revealed that holding soil moisture (i.e., water potential) constant can improve the aboveground biomass (AGB) of C. sativa cultivars grown in living soil. The present study tested two hypotheses, and both were largely supported. The data supports the first hypotheses, which expected AGB to increase with more consistent water pressures throughout the grow cycle. However, this pattern did not hold for one cultivar (Mimosa), which yielded less AGB when irrigated with Blumat systems. Interestingly, the two cultivars that expressed the largest increase in AGB, both had genetics associated with the “Cake” lineage (Birthday Cake & Wedding Cake). Indeed, the fact that different cultivars of plants within the same species can require different propagation and cultivation practices to maximize yield is well documented (Signh and Ghildyal 1980; Wang et al. 2014). This pattern, as should be expected, also holds true for C. sativa, as cultivars have been shown to respond differently to production conditions because of the vast amount of existing genomic diversity within the species from thousands of years of breeding (Backer et al. 2019). Furthermore, genetic variation within selected cultivars of C. sativa have been reported to be more effective at extracting and concentrating heavy metals than others planted simultaneously in the same medium (Higgins and Hussey, unpublished data). Thus, with the vast genetic diversity present in C. sativa, it is important to replicate experiments that focus on a wide range of cultivars to elucidate general patterns in cultivation techniques.
The second hypothesis which expected that soils irrigated with an automated system would provide a more stable soil environment was supported. Over a two-week period, Blumat systems kept soil water potential more stable, closer to the predetermined water pressure target, and did not allow for the large swings between dry and overly wet periods which were observed in beds that were irrigated by hand. Both treatments showed dips below the target irrigation value of 100 mbar on the 12th, 15th, 18th, and 22nd of March. These low mbar values were likely a result of the addition of compost teas to feed the soil microbiology which is standard operating procedure at Gold Leaf Gardens.
While the present study did find a relationship between a consistent soil moisture content and an overall increase of AGB at harvest, the underlying mechanisms were not investigated, and thus provide a clear direction for additional research. Data from previous studies about the interactions between plants and soil water potential can provide insights into likely underlying mechanisms that are working in tandem with one another that could describe the patterns highlighted by the results of the present study. For example, to sustain photosynthesis, water evaporation from the leaves must be supplied from the soil via the soil-to-leaf hydraulic system, and therefore, providing a stable hydrological environment in the soil would allow plants to transpire more efficiently (Hsiao 1973; Enquist et al. 1998; Brodribb 2009; Manzoni et al. 2013; Abdallah et al. 2020). Furthermore, efficient transpiration is also critical to provide nutrients to the leaves, as nutrients are mainly transported with water from the soil to the leaves via the xylem (Yates et al. 2010; Wang et al. 2019).
Perhaps one of the most direct impacts that soil moisture has on plant health is the mechanisms relating to plant transpiration via stomatal guard cell functioning (Saliendra et al. 1995; Mott et al. 1997; Comstock and Mencuccini 1998; Buckley 2005; Hetherington and Woodward 2003; Berr et al. 2010; Buckley 2019). Briefly, the constant availability of moisture in soils allows for increased transpiration rates (Denmead and Shaw 1962; Meerveld and Mcdonnell 2006), which then cascades to permitting maximum photosynthetic potential (Bois et al. 1985) and thus an overall increase in plant growth via an abundance of carbon (C) which can be partitioned to various parts of the plant (Weraduwage et al. 2015). Furthermore, when coupled with elevated levels of CO2 (e.g., 1200 ppm), as is typically implemented in commercial indoor C. sativa facilities, soil moisture content may likely become a limiting factor inhibiting plant growth. Thus, one particular variable that deserves future research would be responses in fan leaf morphometrics (e.g., thickness, stomata size, stomatal density, overall leaf size, etc.) to more consistent soil moisture content, as it would be expected that these plants would display larger leaves as more C is assimilated via increased transpiration and nutrient delivery. Further research should also focus on linking increases in photosynthetic ability, flower development, and ultimately final flower (dry) weight.
In addition to the physiological benefits that constant soil moisture likely has on the growth of C. sativa, another mechanism that likely contributed to the increase in yield is the lack of drought and flood cycles (as is typical for hand waterings; see Fig. 8) promotes a stable environment for the microbiology of the grow medium (Allton and Harris 2007). Soil microbial communities are responsible for the majority of nutrient mineralization and cycling in terrestrial ecosystems (Davidson et al. 1989; Haubensak et al. 2002; Bardgett 2005; Cregger et al. 2012). Their abundance, diversity, community structure and overall activity are directly influenced by a host of abiotic factors, including irrigation (Schadt et al. 2003; Williams and Rice 2007; Collins et al. 2008; Angel 2010; Cregger et al. 2012). Through its impacts on the transport of energy and nutrients, cellular metabolism, osmotic potential, species interactions and as a vector of bacterial motility (Harris 1981; Yancey et al. 1982, Bremer 1999), soil water is considered a key factor responsible for microbial activity and biochemical soil properties (Williams and Rice 2007; Borowik and Wyszkowska 2016). Therefore, the plants exposed to more consistent soil moisture content likely had more nutrient uptake via mass flow and diffusion, which was derived from a more diverse and motile microbial community present in beds fitted with Blumat systems. Indeed, this hypothesis should be tested in future studies.
There are other clear advantages to utilizing automated irrigation systems within commercial facilities aside from the physiological and biological benefits that are provided by a stable soil moisture environment. As the results of the present study showed, hand watering soils have the potential to create repeated underwatered and overwatered periods throughout a grow cycle. A team of experienced gardeners will likely have differing judgement of ideal soil moisture levels, as well as methods for determining what they each consider to be ideal levels. Even if a digital tensiometer is used, keeping soil moisture levels within a stable range is difficult to achieve when the method of water delivery is by hand, and likely will result in the need for multiple labor hours to do so. This becomes increasingly more difficult if production facilities grow multiple cultivars simultaneously (e.g., within the same flower room), as they can differ in their transpiration rates and therefore have different demands on the moisture content of the soil.
It is also important to highlight that some systems are triggered by different metrics. Many automated irrigation systems that facilities deploy to irrigate their soils rely on timers. While this method does likely provide a more consistent soil moisture content relative to hand watering, they largely revolve around anthropogenic variables (e.g., employee schedules, standard operating procedures, and facility workflows), and do not have any direct relationship with the current soil moisture content prior to activating. If the main method of irrigation is determined by time and multiple cultivars share the same grow room, it is plausible that certain genetics are being underwatered, overwatered, or both, as the cultivare requirements can be simply misaligned with facility workflows and irrigation schedules. A likely benefit of using an irrigation system that constantly monitors the water potential of the soil (e.g., Blumat systems), is that it removes the aforementioned sources of human induced error in regard to irrigation. A decrease in both the frequency and intensity of overwatering events would also likely decrease the leaching of important cations, such as potassium (K), and sodium (Na), thus retaining these nutrients within the soil for longer periods of time. Of course, improper irrigation can still occur if the sensor valves are set incorrectly, or the target value (e.g., 100 mbar) is not suitable for the cultivar(s) being grown - another variable in need of investigation.
Conclusions
The data from the present study indicates that utilizing automated irrigation systems can increase AGB of C. sativa. However, genetics of individual cultivars may respond differently when irrigated by different methods, highlighting the need for additional phylogenetic analyses of C. sativa. These data suggest that moving away from hand-watering methods and arbitrary irrigation schedules (which are commonplace in commercial C. sativa cultivation facilities), and in turn adopting irrigation practices relying more on water potential and other soil properties (e.g., soil bulk density, volumetric water content) would provide more robust plants, as well as allow multiple cultivars to simultaneously command their own water requirements, and therefore further close the yield gap. However, exactly how much more stable the soil moisture content is when being irrigated by an automated system that is triggered by changes in soil water potential (e.g., Blumat systems) is when compared to a system that is triggered by timers is yet another direction for future research.
References:
- Abdallah MAB, Durfee N, Mata-Gonzalez R, Ochoa CG, Noller JS (2020) Water use and soil moisture relationships on Western Juniper trees at different growth stages. Water, 12:1596 doi:10.3990/w12061596.
- Allton KE, Harris JA (2007) The effect of microbial communities on soil hydrological processes: A microcosm study utilising simulated rainfall. Geoderma, 142:11-17.
- Angel R, Soares MIM, Ungar ED, Gillor O (2010) Biogeography of soil archaea and bacteria along a steep precipitation gradient. The ISME Journal, 4:553-563.
- Backer R, Schwinghamer T, Rosenbaum P, McCarty V, Eichhorn Bilodeau S, Lyu D, Ahmed MB, Robinson G, Lefsrud M, Wilkins O and Smith DL (2019) Closing the Yield Gap for Cannabis: A Meta-Analysis of Factors Determining Cannabis Yield. Frontiers in Plant Science, 10:495. doi: 10.3389/fpls.2019.00495.
- Bardgett RD. 2005. The biology of soil: a community and ecosystem approach. Oxford University Press, New York, NY
- Bois JF, Cochat PH, Lasceve G (1985) Relationships between transpiration and photosynthesis during a water stress. Acta Horticulture, 171:297-314.
- Borowik A, Wyszkowska (2016) Soil moisture as a factor affecting the microbiological and biochemical activity in soil. Plant Soil Environment, 62:250-255.
- Bremer E (1999) Microbial stress responses to high-osmolality environments: uptake and synthesis of compatible solutes. American Zoologist, 39:6a.
- Brodribb TJ (2009) Xylem hydraulic physiology: the functional backbone of terrestrial plant productivity. Plant Science, 177:245–251.
- Buckley TN (2005) The control of stomata by water balance. New Phytologist, 168:275-292.
- Buckley TN (2019) How do stomata respond to water status? New Phytologist, 224:21-36.
- Burstein, S (2015) Cannabidiol (CBD) and its analogs: a review of their effects on inflammation. Bioorganic and Medical Chemistry, 23:1377–1385. doi: 10.1016/j.bmc.2015.01.059
- Chandra S, Lata H, Khan IA, Elsohly MA (2011a) Photosynthetic response of Cannabis sativa L., an important medicinal plant, to elevated levels of CO2. Physiology and Molecular Biology of Plants, 17:291–295. doi: 10.1007/s12298-011- 0066-6
- Chandra S, Lata H, Khan IA, Elsohly MA (2011b) Temperature response of photosynthesis in different drug and fiber varieties of Cannabis sativa L. Physiology and Molecular Biology of Plants, 17:297–303. doi: 10.1007/s12298-011-0068-4
- Chandra, S., Lata, H., Mehmedic, Z., Khan, I. A., and Elsohly, M. A. (2015) Light dependence of photosynthesis and water vapor exchange characteristics in different high Δ9-THC yielding varieties of Cannabis sativa L. Journal of Applied Research on Medicinal and Aromatic Plants, 2:39–47. doi: 10.1016/j.jarmap.2015.03.002
- Collins SL, Sinsabaugh RL, Crenshaw C, Green L, Porras-Alfaro A, Stursova M, Zeglin (2008) Pulse dynamics and microbial processes in aridland ecosystems. Journal of Ecology, 96:413-420.
- Comstock JP, Mencuccini M (1998) Control of stomatal conductance by leaf water potential in Hymenoclea salsola (T. & G.), a desert subshrub. Plant, Cell & Environment, 21:1029-1038.
- Cregger MA, Schadt CW, McDowell NG, Pockman WT, Classen AT (2012) Response of the soil microbial community to changes in precipitation in a semiarid ecosystem. Applied and Environmental Microbiology, 78:8587-8594.
- Davidson EA, Eckert RW, Hart SC, Firestone MK (1989) Direct extrac- tion of microbial biomass nitrogen from forest and grassland soils of Cal- ifornia. Soil Biology and Biochemistry, 21:773–779.
- Denmead OT, Shaw RH (1962) Availability of Soil Water to Plants as Affected by Soil Moisture Content and Meteorological Conditions. Agronomy Journal, 5:385-390.
- Giacoppo S, Mandolino G, Galuppo M, Bramanti P, Mazzon E (2014) Cannabinoids: new promising agents in the treatment of neurological diseases. Molecules, 19:18781–18816. doi: 10.3390/molecules191118781.
- Harris R (1981) In: Parr JF, Gardner WR, Elliott LF (Eds.), Effect of Water Potential on Microbial Growth and Activity in Soils in Water Potential Relations in Soil Microbiology. Soil Science Society of America Journal, Madison, WI, pp. 23–96.
- Haubensak KA, Hart SC, Stark JM (2002) Influences of chloroform exposure time and soil water content on C and N release in forest soils. Soil Biology and Biochemistry, 34:1549 –1562.
- Hetherington AM, Woodward FI (2003) The role of stomata in sensing and driving environmental change. Nature, 424:901-908.
- Hill AJ, Williams, CM, Whalley BJ, Stephens GJ (2012) Phytocannabinoids as novel therapeutic agents in CNS disorders. Pharmacology and Therapeutics, 133:79–97. doi: 10.1016/j.pharmthera.2011.09.002.
- Hsiao TC (1973) Plant responses to water stress. Annual Review of Plant Physiology and Plant Molecular Biology, 24:519–570.
- Lydon J, Teramura AH, Coffman CB (1987) UV-B radiation effects on photosynthesis and cannabinoid production of two Cannabis sativa chemotypes. Photochemistry and Photobiology, 46:201–206. doi: 10.1111/j.1751-1097.1987.tb04757.x.
- Marti G, Schnee S, Andrey Y, Simoes-Pires C, Carrupt PA, Wolfender JL, et al (2014) Study of leaf metabolome modifications induced by UV- C radiations in representative Vitis, Cissus and Cannabis species by LC- MS based metabolomics and antioxidant assays. Molecules, 19:14004–14021. doi: 10.3390/molecules190914004.
- Manzoni S, Vico G, Katul G, Porporato A (2013) Biological constraints on water transport in the soil–plant–atmosphere system. Advances in Water Resources, 51:292–304. doi:10.1016/j.advwatres.2012.1003.1016.
- Mott KA, Denne F, Powell J (1997) Interactions among stomatal in response to perturbations in humidity. Plant, Cell & Environment, 20:1098-1107.
- Meerveld IV, Mcdonnell J (2006) On the interrelationships between topography, soil depth, soil moisture, transpiration rates and species distribution at the hillslope scale. Advances in Water Resources, 29:292-310.
- Kim SY, Lee SH, Freeman C, Fenner N, Kang H (2008) Comparative analysis of soil microbial communities and their responses to the short-term drought in bog, fen, and riparian wetlands. Soil Biology and Biochemistry, 40:2874–2880.
- Saliendra NZ, Sperry JS, Comstock JP (1995) Influence of leaf water status on stomatal response to humidity, hydraulic conductance, and soil drought in Betula occidentalis. Planta, 196:357-366.
- Schadt CW, Martin AP, Lipson DA, Schmidt SK (2003) Seasonal dynamics of previously unknown fungal lineages in tundra soils. Science, 301:1359 –1361.
- Sign R, Ghildyal BP (1980) Soil submergence on Nutrient uptake, growth and yield of five corn cultivars. Agronomy Journal, 72:737:741.
- Silva CC, Guido ML, Ceballos JM, Marsch R, Dendooven L (2008) Production of carbon dioxide and nitrous oxide in alkaline saline soil of Texcoco at different water contents amended with urea: A laboratory study. Soil Biology and Biochemistry, 40:1813–1822.
- Unger IM, Kennedy AC, Muzika RM (2009) Flooding effects on soil microbial communities. Applied Soil Ecology, 42: 1–8.
- Volkow ND, Baler RD, Compton WM, Weiss SR (2014) Adverse health effects of marijuana use. New England Journal of Medicine, 370,2219–2227. Doi: 10.1056/NEJMra1402309.
- Walker TS, Bais HP, Grotewold E, Vivanco JM (2003) Root exudation and rhizosphere biology. Plant Physiology, 132:44–51.
- Wang C, He J, Zhao T-H, Cao Y, Wang G, Sun B, Yan X, Guo W and Li M-H (2019) The Smaller the Leaf Is, the Faster the Leaf Water Loses in a Temperate Forest. Frontiers in Plant Science, 10:58. doi: 10.3389/fpls.2019.00058.
- Weraduwage SM, Chen J, Anozie FC, Morales A, Weise SE, Sharkey TD (2015) The relationship between leaf area growth and biomass accumulation in Arabidopsis thaliana. Plant Science, 6:167. doi:10.3389/fpls.2015.00167
- Williams MA, Rice CW (2007) Seven years of enhanced water availability influence the physiological, structural, and functional attributes of a soil microbial community. Applied Soil Ecology 35:535-545.
- Winston ME, Hampton-Marcell J, Zarraonaindia I, Owens SM, Moreau CS, Gilbert JA, et al. (2014).Understanding cultivar-specificity and soil determinants of the cannabis microbiome. PLoS ONE 9:e99641. doi: 10.1371/journal.pone.0099641.
- Borowik A, Wyszkowska J (2016) Soil moisture as a factor of affecting the microbiological and biochemical activity of soil. Plant Soil and Environment, 62:250-225.
- Yancey PH, Clark ME, Hand SC, Bowlus RD, Somero GN (1982) Living with water-stress—evolution of osmolyte systems. Science, 217:1214–1222.
- Yates MJ, Anthony Verboom G, Rebelo AG, Cramer MD (2010) Ecophysiological significance of leaf size variation in proteaceae from the cape floristic region. Functional Ecology, 24:485–492. Doi: 10.1111/j.1365-2435.2009. 01678.x.
Acknowledgements
The authors would like to thank Nathan Gibbs and Gavin Wada of Gold Leaf Gardens for the useage of their facility and plants for the study. Bryant Mason provided valuable feedback during the preperation of the manuscript.
Funding
No external funding sources contributed to the present study.
Author information
Affiliations
KIS Organics
Ben Higgins, PhD & Tad Hussey
Gold Leaf Gardens
Ben Higgins, PhD
Contributions
Tad Hussey and Ben Higgins contributed equally to the development of the experimental design of the study. All data was collected and analyzed by Ben Higgins. Manuscript preperation was competed by Ben Higgins, and edited by Tad Hussey. The authors read and approved the final draft of the manuscript prior to submission.
Corresponding author
Corresponence to Ben Higgins







